-
Par trichard le 16 Octobre 2006 à 15:10
http://www.snv.jussieu.fr/vie/bib/dos-doc/1documents.htm
Modélisation du concept de "boîte noire" :

Modélisation cybernétique d'un système effecteur :

Modélisation cybernétique d'un régulateur en
constance. Il est caractérisé par une rétroaction
négative :
Modélisation cybernétique d'un régulateur en
tendance. Il est caractérisé par une rétroaction
positive :
Modélisation cybernétique d'un homéostat. Noter
l'intervention de deux systèmes, un 'réglant' et un 'réglé'.
Grâce à une voie de communication, le système réglant
peut agir sur les grandeurs d'entrée du système réglé,
et ainsi positionner la grandeur réglée au niveau de la valeur
de consigne :
En cybernétique, un homéostat peut être décrit d'une manière un peu plus détaillée, par l'association de deux systèmes :
* Un système réglé : le compartiment dans lequel est définie la grandeur réglée, caractérisé en particulier par ses grandeurs d'entrée (paramètres affectant la grandeur réglée) et par sa grandeur de sortie (la grandeur réglée).
* Un système réglant : agit en permanence sur le système réglé, afin de maintenir la grandeur réglée aussi proche que possible de la valeur de consigne. Il est composé de trois éléments :
1. Un émetteur, ou émetteur-capteur : détecte les variations de la grandeur réglée par rapport à la grandeur de consigne, et traduit cet écart en une information codée, transmise par le transmetteur.
2. Un transmetteur : transmet l'information codée par l'émetteur au récepteur de la voie de communication.
3. Un récepteur : c'est l'effecteur du système réglant. Il traduit le message codé qu'il reçoit en une action capable de rectifier la valeur de la grandeur réglée.Dans la très grande majorité des cas, les paramètres physiologiques
du milieu intérieur d'un organisme tel que l'Homme sont maintenu constants
: il s'agit d'homéostats, fonctionnant grâce à des régulateurs
en constance. Toutefois, dans certains cas, des régulateurs en tendance
peuvent intervenir : ils résultent de la modification, de manière
active, de la grandeur réglée, dans le sens d'une diminution ou
d'une augmentation. Toutefois, ces modifications ne sont pas infinies et sont
limitées dans le temps : elles sont la conséquence du déplacement
transitoire de la valeur de la grandeur de consigne vers une nouvelle valeur
bien déterminée.Dans ces phénomènes, des servomécanismes interviennent
: il s'agit de dispositifs permettant de modifier la valeur de la grandeur de
consigne. Ils mettent en jeu des régulateurs en tendance pour modifier
la grandeur réglée, et des régulateurs en constance pour
la maintenir ensuite à la nouvelle grandeur de consigne. Il s'agit donc
ici d'une notion de pilotage : le servomécanisme permet
une variation précise, commandé par un niveau d'organisation supérieur
de l'organisme. votre commentaire
votre commentaire
-
Par trichard le 12 Octobre 2006 à 22:51
http://www.inrp.fr/Acces/biotic/procreat/amp/html/GametesMurs.htm
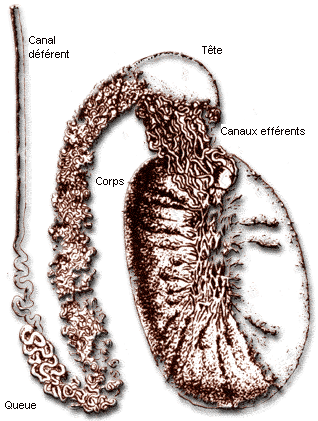
Représentation de l'épididyme et de ses
relations avec le testicule
La tête de l'épididyme (en haut) est assez volumineuse,
le corps de l'épididyme longe le testicule et la queue de l'épididyme
est la zone où les spermatozoides son stockés.
D'après http://spermiologie.u-strasbg.fr votre commentaire
-
Par trichard le 12 Octobre 2006 à 16:31
http://www.snv.jussieu.fr/vie/bib/dos-doc/1documents.htm
Première étape : formation d'une ébauche de gonade indifférenciée
crête génitale et cellules germinales
Les cellules germinales migrent, chez tous les Vertébrés,
pour aller coloniser la crête génitale. Chez les Mammifères,
ces cellules sont originaire du mésoderme extra-embryonnaire. Elles
accomplissent leur migration en passant par l'allantoïde :
Deuxième
étape : formation d'une gonade
développement de la crête génitale
Première étape : développement identique
chez l'homme et la femmeA. A 4 semaines de développement, la crête génitale
est visible. Elle comporte les cellules germinales. Elle se développe
à côté de la crête mésonéphrétique,
qui comporte le futur canal de Wolff.
B. Après 6 semaines de développement, le canal
de Müller est apparu dans la crête mésonéphrétique.
Les cordons sexuelles primitifs se développent à partir
de l'épithelium de la crête génitale.Deuxième étape : développement différent
chez l'homme et la femmeC. Au stade 8 semaines, les cordons
sexuels, pour les foetus de type mâle ou XY, vont se développer
en pénétrant dans le tissu conjonctif et former un réseau
relié au niveau interne par les cordons du rete
testis. L'extrémité de
l'ensemble des cordons sexuels va se détacher de l'épithélium
de la crête génitale. Ces cordons seront séparés
par une matrice extracellulaire, l'albuginée.
D. Au stade 16 semaines, les cordons
sexuels ou testiculaires contenant des cellules germinales (précurseurs
des gamètes) se développent , augmentant alors la taille
du testicule. Le réseau formé par les cordons testiculaires
et les cordons du rete testis
est relié au canal déférent (tube dans lequel les
spermatozoides passent dans l'urètre et sortent de l'organisme)
par des restes de tube mésonéphrétique (canaux
efférents).E. Au stade 8 semaines, les cordons sexuels primitifs dégénérent
; seul l'épithélium de surface reste et produit des cordons
corticaux .F. Au stade 20 semaines, la folliculogénése,
c'est-à-dire la formation des follicules, démarre. On
observe une migration des follicules entourant l'ovogonie dans la crête
génitale mais il n'y a pas de connections avec le canal de Müller.Pour les foetus XY, on observe dans la crête
génitale la formation de deux types de cordons : les cordons testiculaires
qui contiennent les cellules germinales qui produiront les futurs spermatozoïdes
; et les cordons du rete testis qui se trouvent à l'extrémité
des cordons testiculaires. Le canal de Wolff est relié aux cordons
du rete testis par des restes du tube mésonéphrétique
et se différencie en canal déférent pour permettre
la sortie des spermatozoïdes. Au cours de ce développement,
les cellules du mésenchyme interstitiel des testicules vont devenir
les cellules de Leydig (production de la testostérone qui favorise
le maintien du canal de Wolff) et les cellules des cordons testiculaires,
autres que les cellules germinales, vont se différencier en cellules
de Sertoli (nutrition des spermatozoïdes et sécrétion
de l'hormone anti-Müllerienne qui favorise la dégénérescence
du canal de Müller). Pendant la puberté, il y a formation des
tubes séminifères par creusement des cordons testiculaires,
et les cellules germinales produisent les spermatozoides.Chez les foetus de type femelle ou XX, les cordons
sexuels primitifs dégénérent. Néanmoins, l'épithélium
de surface produit de nouveaux cordons qui ne pénétrent
pas dans le tissu conjonctif mais qui restent en contact avec la surface
corticale de la crête génitale. Ils forment les amas cellulaires
différenciés de la granulosa (d'origine somatique) et entourent
les cellules germinales. Les cellules des thèques (cellules périphériques
et protectrices) se forment ensuite autour de chaque ensemble (granulosa
+ cellule germinale), pour former les follicules. Ces
follicules sécrétent des hormones stéroïdes.
La formation d'un follicule n'est possible que s'il entoure une cellule
germinale. Pour les individus XX, on observe une dégénérescence
du canal de Wolff par l'absence de testostérone et le canal de
Müller se développe pour former l'appareil génital
femelle (l'oviducte, l'utérus,le canal cervical et le vagin supérieur).
Troisième étape
: acquisition du sexe phénotypiqueLes gonades en développement secrètent un certain nombre d'hormones. Ces hormones permettent le développement de l'ensemble de l'appareil
génital vers un phénotype mâle ou femelle. Cette acquisition
du sexe phénotypique est visible en particulier au niveau des canaux
de Wolff et de Müller : Au stade indifférencié de l'appareil génital,
les canaux de Wolff et de Müller sont présents.Le testicule secrète testostérone et hormone anti-Müllerienne.
Les canaux de Wolff constituent le spermiducte chez le
mâle des vertébrés. Ils sont en relation avec le mésonéphros
mis en place au cours de la quatriéme semaine du développement.
L'extension du canal de Wolff jusqu'à l'urètre se fait pendant
la cinquème semaine.Ces canaux dégénérent chez
la femelle. Les canaux de Müller se développent en parallèle
aux canaux de Wolff par une invagination de l'épithélium au niveau
du pronéphros (région antérieure du mésonéphros)
au cours de la sixième semaine. Ils deviendront les oviductes de la femelle
et déboucheront dans l'utérus. Ces canaux dégénérent
chez le mâle. votre commentaire
-
Par trichard le 11 Octobre 2006 à 17:00
http://www.snv.jussieu.fr/vie/conferences/morphovg_ai_01/03localisation/images/soja16-480.mov
http://www.snv.jussieu.fr/vie/dossiers/plantules/soja/gravi-320.mov
http://www.snv.jussieu.fr/vie/conferences/morphovg_ai_01/02plantules/images/barquette-640.mov
http://www.snv.jussieu.fr/bmedia/racine/images/camera.gif
votre commentaire
-
Par trichard le 11 Octobre 2006 à 15:45
http://www.ac-nantes.fr:8080/peda/disc/svt/soja/TPsoja.htm
Les mécanismes
de l'action de l'éthylène ne sont pas encore complètement
identifiés. Actuellement, on sait que l'éthylène se fixe
sur un récepteur membranaire. Les études sur Arabidopsis ont
permis de déterminer que cette fixation déclenche la succession
d'évènements ci-dessous : votre commentaire
votre commentaire Suivre le flux RSS des articles de cette rubrique Suivre le flux RSS des commentaires de cette rubrique
Suivre le flux RSS des articles de cette rubrique Suivre le flux RSS des commentaires de cette rubrique
Mon petit cahier de sciences naturelles
-

Platynereis dumerili




{kind=link}
{kind=link}