-
http://www.larecherche.fr/content/actualite-sapiens/article?id=30599

Pour ce nouveau numéro des Dossiers de La Recherche, en kiosque aujourd'hui, nous vous proposons de plonger dans le passé, sur les traces de l'homme de Neandertal. Découvrez comment la génétique se joint à la paléontologie et l'archéologie pour apporter de nouveaux éclairages sur notre lointain ancêtre.Comment retracer l'histoire de Neandertal ? Si l'on est loin de pouvoir le faire entièrement, on connaît un peu mieux son parcours. Il s'est métissé à faible dose avec l'homme moderne. Il partageait une grande partie de son génome avec une nouvelle lignée humaine révélée par son ADN, l'homme de Denisova.
Ces récentes découvertes sont dues au progrès d'une jeune discipline, la paléogénétique. Vont-elles nous conduire à revoir notre vision de la préhistoire ? D'ores et déjà, avec les nouveaux fossiles trouvés en Asie, l'épopée d'Homo sapiens apparaît plus complexe qu'on ne le pensait. Et l'on revisite les scénarios de la disparition de Neandertal.
 votre commentaire
votre commentaire
-
La théorie de l'évolution est en évolution : des termes à proscrire, d'autres à inscrire :
-
du scalisme à la cladistique
-
de l'invertébré à la clade
-
du chaînon manquant à l'ancêtre commun
-
du fossile vivant au fossile tout court
-
de l'arbre généalogique à l'arbre phylogénétique
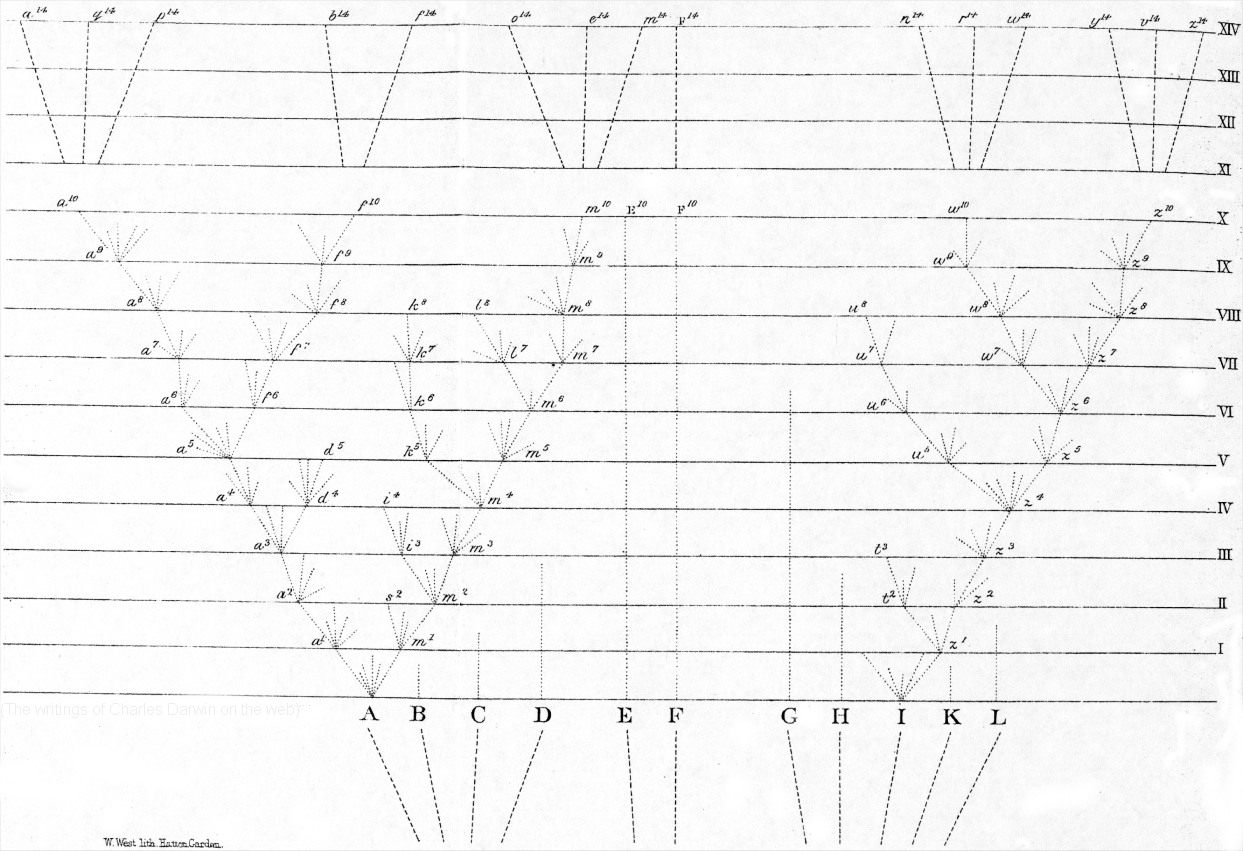
La cladistique a rendu caduque la recherche de l’« ancêtre commun ». Certes, toute espèce actuelle ou fossile a des ancêtres plus ou moins lointains. L’Homme et le Chimpanzé ne font pas exception à cette règle ; ils ont eu un ancêtre commun récemment (à l’échelle des temps géologiques). Cependant, dans la mesure où, comme le soulignait Darwin, les descendants sont forcément différents de leurs ancêtres (ceci est le sens même de l’évolution), il n’y a pas de moyen d’identifier à coup sûr cet ancêtre sur des critères descriptifs parmi les fossiles retrouvés. Un arbre phylogénétique ne peut pas illustrer les processus évolutifs eux-mêmes. De plus, l’évolution n’est pas graduelle, mais fait intervenir des ruptures, des discontinuités.
Exercice (II.2 Liban 2010) : phylogénie des Téthythériens
§ l'Humain a été situé dans le groupe des vertébrés, le chapitre suivant sera consacré à la place de l'homme dans l'évolution.
33 - La place de l’Homme dans le règne animal, la lignée humaine
Film cnrs : http://www.cnrs.fr/biodiversite2010/spip.php?article5
choisissez dans la liste : "primates, homologie et parenté"
331 - Place de l'humain au sein du vivant
données – notamment moléculaires – sur les primates : voir tableau photocopié des gènes HCG et HLP, et celui des globines
parenté étroite entre le Chimpanzé et l’Homme : 98% de gènes communs
remonter l'arbre du vivant à partir d'Homo sapiens : http://tolweb.org/Homo_sapiens/16421#titlefigcaption
classification simplifiée : http://www.pedagogie.ac-nantes.fr/html/peda/svt/classifs/terminale/htm/terminale.htm
votre commentaire
-
-
Les portions codantes de l’ADN comportent l’information nécessaire à la synthèse de chaînes protéiques issues de l’assemblage d’acides aminés.
Chez les eucaryotes, la transcription est la fabrication, dans le noyau, d’une molécule d’ARN pré-messager, complémentaire du brin codant de l’ADN. Après une éventuelle maturation, l’ARN messager est traduit en protéines dans le cytoplasme.
Un même ARN pré-messager peut subir, suivant le contexte, des maturations différentes et donc être à l’origine de plusieurs protéines différentes.
La séquence des nucléotides d’une molécule d’ADN représente une information. Le code génétique est le système de correspondance mis en jeu lors de la traduction de cette information. À quelques exceptions près, il est commun à tous les êtres vivants.
§ comment une information génétique traduit-elle au niveau d'un ensemble cellulaire, l'organisme ? De la population ?
13- Des gènes à la réalisation des phénotypes
131- Un caractère à différentes échelles
A1/ Biochimie des groupes sanguins
Manuel p67 : groupes sanguins
au niveau micro et moléculaire : http://www.snv.jussieu.fr/vie/dossiers/gpes-sanguins/index.htm
différentes échelles d’un phénotype
expression génétique, protéine, facteurs d'expression, variabilité
A2/ Cas de l'albinisme
Manuel p66 : albinisme chaîne de biosynthèse moléculaire
Tyr - [tyrosinase] → dopaquinone… → mélanine
A3/ Cas de la drépanocytose
Diap drepanocytose
manuel p62 à 65
Phénotype mol, micro, macroscopique,
132- Gènes et environnement
A1/ Comparaison de gènes sur banque de séquences / logiciel anagène
Manuel p 64
drépanocytose : fichier → thèmes d'étude → relation pheno-génotype → phénotype drépanocytaire → comparer les séquences → repérer les différences
albinisme : fichier → banque de séquences → tyrosinase → tyrcod1 & tyralba1 → comparer
groupes sanguins : fichier → banque de séquences →
gène, allèles, génotype
votre commentaire
-
http://www.larecherche.fr/content/actualite-sapiens/article?id=30744

Les circuits neuronaux permettant de percevoir la douleur commencent à se former deux semaines avant la naissance, selon des neurologues britanniques.Longtemps, on a pensé que le nouveau-né ne ressentait pas la douleur, parce que son cerveau n’était pas assez mature. Jusque dans les années 1960 l’anesthésie des nouveau-nés pour certaines interventions médicales était même jugée inutile. Depuis, plusieurs études ont prouvé que les bébés sont sensibles à la douleur dès leur première semaine de vie. Et les pratiques médicales ont changé en conséquence.
Une étude britannique va encore plus loin : elle démontre que les réseaux neuronaux permettant de distinguer la douleur d’une stimulation tactile « normale » commencent à émerger vers 35 semaines de grossesse, soit deux semaines avant la naissance à terme [1] .
Lorenzo Fabrizi, de l’université de Londres, et ses collègues ont sélectionné 46 bébés : 25 bébés nés à terme, entre 37 et 41 semaines de grossesse, et 21 bébés nés prématurément entre 28 et 36 semaines de grossesse. Ils ont réalisé un électroencéphalogramme de ces bébés afin d’enregistrer l’activité de leur cerveau lors de deux stimulations tactiles différentes.
La première stimulation, non douloureuse, consistait en un toucher du talon avec un marteau à réflexe. La seconde, douloureuse, était une piqûre sur le talon couramment pratiquée chez les nouveau-nés pour effectuer des prélèvements sanguins. Les neurologues ont constaté que selon l’âge des bébés la réponse cérébrale à ces deux stimulations variait.
Stimulation électrique.
Chez la plupart des bébés nés à terme, chaque stimulation a provoqué une activité cérébrale spécifique, preuve que les circuits neuronaux propres au toucher et à la douleur s’étaient différenciés. Au contraire, chez les prématurés, les deux stimulations ont provoqué le plus souvent les mêmes bouffées d’activité neuronale, autrement dit une réponse cérébrale non spécifique.
Cependant, dans 7 % des cas, une activité spécifique a été enregistrée en réponse à la stimulation tactile, et cette proportion a été de 33 % pour la stimulation douloureuse. Surtout, les neurologues ont constaté que la probabilité que le cerveau produise une réponse spécifique augmentait avec l’âge des bébés, alors que la probabilité qu’il produise des bouffées non spécifiques diminuait : les proportions commencent à s’inverser vers 35 semaines de grossesse.
Ces résultats suggèrent que les circuits neuronaux permettant de distinguer une stimulation douloureuse d’une stimulation tactile normale se mettent en place deux semaines avant la naissance.
« En 2010, cette équipe avait montré que, chez les bébés nés à 36 semaines de grossesse, une stimulation douloureuse provoque une réponse neuronale spécifique, précise Édouard Gentaz, de l’université de Grenoble. Cette nouvelle étude, qui porte sur des bébés nés à des âges très différents, apporte des résultats plus précis, en montrant que la signature neuronale de la douleur émerge entre 35 et 37 semaines de grossesse. Ces résultats encouragent à être encore plus vigilants sur la nature des soins et le contrôle de l’environnement, stressant, des services de néonatalogie : un défaut de prise en charge de la douleur chez les prématurés pourrait avoir un impact à long terme sur leur développement . »
Jacques Abadie
Vous êtes sur Sapiens > article
[1] L. Fabrizi et al., Current Biol., 2010. votre commentaire
-
323 – Apport des données moléculaires
© matrices-arbres
arbre humain-chimp-grenouille-chauve souris
les données moléculaire confirment les données macroscopiques : états dérivés => parenté => ancêtre commun.
Plus des espèces partagent d'états dérivés, ou plus leur degré de similitude moléculaire est important, plus elles sont proches et leur dernier ancêtre commun est récent.
L'ancêtre commun à deux espèces présente les états dérivés de ses ascendants. Une partie de la population de ces ascendants possède des innovations génétiques qui sont transmises aux générations suivantes jusqu'à différentiation d'une nouvelle espèce.
324 – Apport des données paléontologiques
© matrices-arbres
arbre tetrapodes
ancêtre commun = espèce et non individu, entité hypothétique
espèce fossile exprime des états dérivés, ce n'est donc pas un "chaînon manquant" ou un "ancêtre commun", on ne le place pas sur un noeud mais sur une branche de l'arbre phylogénétique.
Conclusion
Diap évolution de « l'arbre du vivant » au cours de l'histoire des sciences :
http://darwin-online.org.uk/converted/published/1872_Origin_F391/1872_Origin_F391_figdiagram.jpg
http://upload.wikimedia.org/wikipedia/commons/archive/9/9c/20090615174705!Darwin_ape.jpg
http://s1.e-monsite.com/2008/08/31/86429592qui-a-vu-chantal-jpg.jpg
http://syl20.jeblog.fr/le-chainon-manquant-a644142#comments
http://roughguidetoevolution.blogspot.com/2008_10_01_archive.html
http://hebergement.ac-poitiers.fr/ecoles79/inspections/thouars/classif/dossier1.HTM
La théorie de l'évolution est en évolution : des termes à proscrire, d'autres à inscrire :
-
du scalisme à la cladistique
-
de l'invertébré à la clade
-
du chaînon manquant à l'ancêtre commun
-
du fossile vivant au fossile tout court
-
de l'arbre généalogique à l'arbre phylogénétique
La cladistique a rendu caduque la recherche de l’« ancêtre commun ». Certes, toute espèce actuelle ou fossile a des ancêtres plus ou moins lointains. L’Homme et le Chimpanzé ne font pas exception à cette règle ; ils ont eu un ancêtre commun récemment (à l’échelle des temps géologiques). Cependant, dans la mesure où, comme le soulignait Darwin, les descendants sont forcément différents de leurs ancêtres (ceci est le sens même de l’évolution), il n’y a pas de moyen d’identifier à coup sûr cet ancêtre sur des critères descriptifs parmi les fossiles retrouvés. Un arbre phylogénétique ne peut pas illustrer les processus évolutifs eux-mêmes car l’évolution n’est pas graduelle, mais fait intervenir des ruptures, des discontinuités.
votre commentaire
-
Mon petit cahier de sciences naturelles
-

Platynereis dumerili




{kind=link}
{kind=link}
{kind=link}
{kind=link}